Chapter 2: Light and the Eye
2.3 The Retinal Network
As we have already seen, the retina is composed of several layers and contains the photoreceptors, the specialized receptor cells for the initial processing of visual stimuli. The photoreceptors (rods and cones) contain visual pigments that absorb light which causes a change in their membrane potentials. This light transduction happens in the outer segments of the rods and cones (see Figure 2.5). When light (of the right wavelength) hits 11-cis retinal, it changes shape (to all-trans form), acting as a switch to start an enzyme cascade in the cell, which eventually changes the rate at which photoreceptors release neurotransmitters (see Figure 2.6).
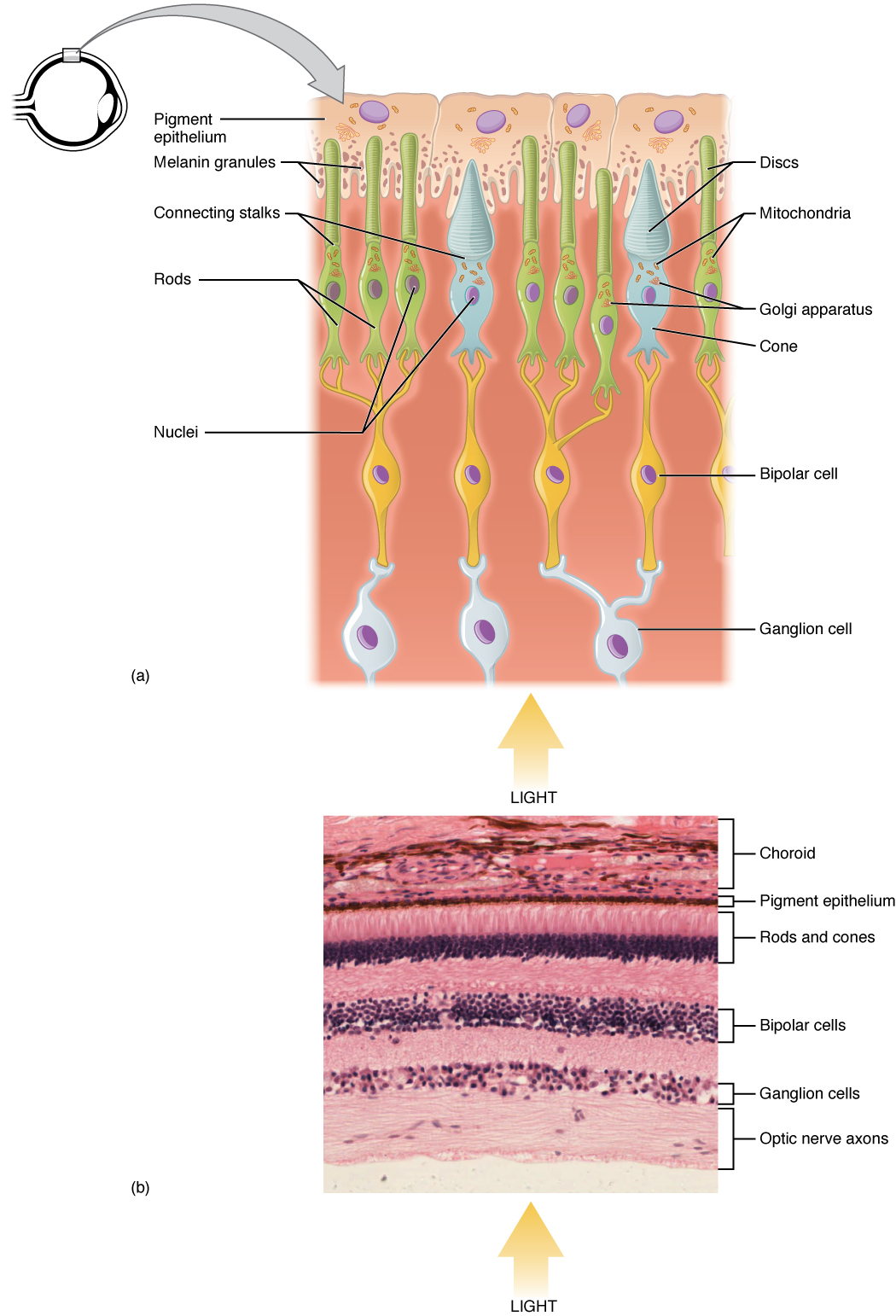
As we will see in the next module, after absorbing light, all-trans retinal needs to be turned back into 11-cis retinal so that it can be recycled. So it breaks off its opsin, finds its way to the pigment epithelium, gets bent back into shape, and finds its way back to an opsin. This is why photoreceptor outer segments need to be close to the pigment epithelium, which is the back layer behind the retina where visual pigments are replenished.
The change in membrane potential caused by light transduction alters the amount of neurotransmitter that the photoreceptor cells release onto bipolar cells. The bipolar cells in turn send neural signals to the ganglion cells. The axons of the ganglion cells make up the optic nerve and transmits information from the eye to the brain. Rods and cones behave differently to each other. Rods are very sensitive to light and help us to see in the dark, however, they are not good at discerning details and they don’t permit us to see in color. In contrast, there are three types of cones, which are activated during daylight and help us to see fine details and colors.
Synaptic Convergence of Photoreceptors
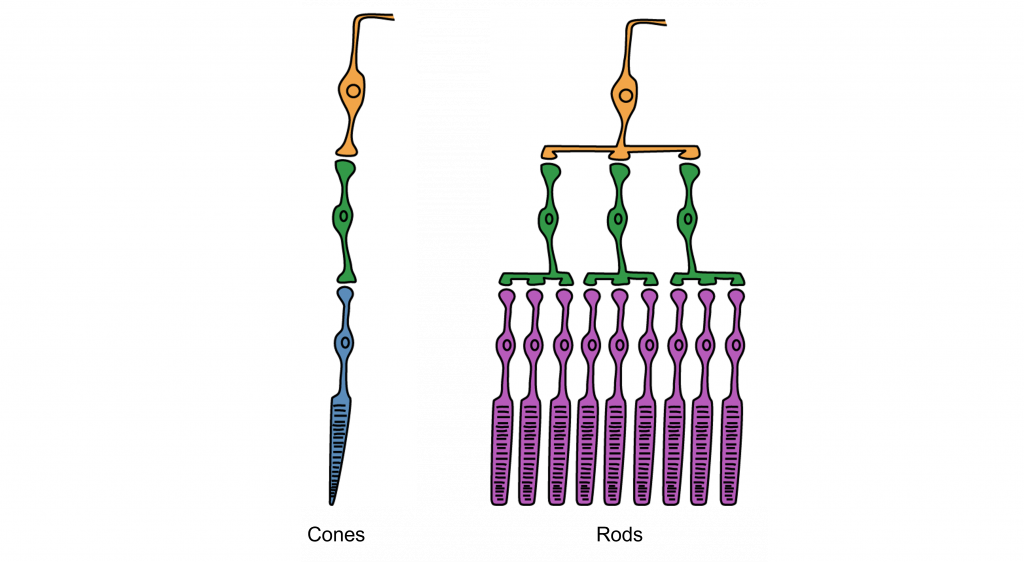
At the fovea, each cone communicates directly with its own bipolar cell. However, for the receptors in other parts of the retina, there is synaptic convergence, where multiple photoreceptors “feed” a single bipolar cell (see Figure 2.7). There are many more rods in the retina than cones and so not surprisingly, there is more convergence for rods than cones. An advantage of having these high-convergence network is the ability to add many small signals together to create a larger signal that meets the threshold of excitation for a bipolar cell. This is why we are able to see very dim stars at night when we are using our rod or scotopic vision. However, a disadvantage of this type of organization is that it is difficult to identify exactly which photoreceptor is activated by the incoming light, which is why accuracy is poor when seeing stimuli in our peripheral vision. This is one of the reasons that we cannot actually read text in our peripheral vision or when there is not much light in the environment.
In general, cone cells have lower synaptic convergence than rods. However, because of this low-convergence organization, cone cells are highly effective at precisely identifying the location of incoming light, especially at the fovea, where there is no convergence. The separation of the cones at the fovea determines our visual acuity — i.e., the smallest detail that we can see. For most adults, this is 1 minute of arc and corresponds to the 20/20 line on the eye chart. However, we are unable to detect very low levels of light using our cone vision.
OpenStax, Anatomy and Physiology Chapter 14.1 Sensory Perception
Provided by: Rice University.
Access for free at https://openstax.org/books/anatomy-and-physiology/pages/14-1-sensory-perception
License: CC-BY 4.0
Adapted by: Kathryn TaterkaPsychology by Jeffrey C. Levy, VisionProvided by: B.C. Faculty Pressbooks
URL: https://pressbooks.bccampus.ca/thescienceofhumanpotential/chapter/vision/
License: CC BY 4.0
Adapted by: Kathryn TaterkaConvergence is adapted from Introduction to Neuroscience Copyright © 2022 by Valerie Hedges is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
